|
||||
|
|
Глава 7. Наследование формы 7.1. Генетика и наследственность Наследственные различия между организмами, одинаковыми в других отношениях, зависят от генетических различий; эти последние зависят от различий в структуре ДНК или в ее расположении в хромосомах, а эти различия ведут к изменениям в структуре белков или к изменениям в контроле синтеза белка. Эти фундаментальные открытия, подтвержденные множеством тщательно поставленных экспериментов, дают достаточно ясное понимание процесса наследования белков, а также характеристик, которые более или менее непосредственно зависят от определенных белков, например, серповидно-клеточная анемия и наследственные дефекты метаболизма. Напротив, наследственные различия формы обычно не имеют непосредственной и очевидной связи с изменениями структуры или синтеза определенных белков. Тем не менее такие изменения могут влиять на морфогенез различными способами, через воздействие на метаболические ферменты, ферменты синтеза гормонов, структурные белки, белки в клеточных мембранах и так далее. Много примеров такого влияния уже известно. Но если различные химические изменения приводят к изменениям или искажениям нормальной картины морфогенеза, то что же определяет саму эту нормальную картину? Согласно механистической теории, клетки, ткани, органы и организмы принимают соответствующие формы в результате синтеза нужных химических компонентов в нужных местах в нужное время. Предполагается, что морфогенез осуществляется самопроизвольно вследствие сложных физико-химических взаимодействий в соответствии с законами физики. Но с какими законами физики? Механистическая теория просто оставляет этот вопрос открытым (раздел 2.2). Гипотеза формативной причинности предлагает новый ответ на этот вопрос. До тех пор пока она дает интерпретацию биологического морфогенеза, которая подчеркивает аналогию с физическими процессами, такими как кристаллизация, а также приписывает важную роль энергетически неопределенным флуктуациям, она, скорее, выполняет, нежели отрицает ожидания механистической теории. Но если последняя приписывает практически все феномены наследственности генетическому наследованию, воплощенному в ДНК, то гипотеза формативной причинности предполагает, что организмы наследуют также морфогенетические поля прошлых организмов того же вида. Этот второй тип наследования осуществляется путем морфического резонанса, а не через гены. Таким образом, наследственность здесь включает как генетическое наследование, так и морфический резонанс от прошлых подобных форм. Рассмотрим следующую аналогию. Музыка, льющаяся из громкоговорителя радиоприемника, зависит как от материальных структур приемника и от энергии, приводящей его в действие, так и от передачи, на которую настроен радиоприемник. Конечно, на музыку могут влиять изменения в проводах, транзисторах, конденсаторах и так далее, и она прекращается, когда вынимается батарейка. Тот, кто ничего не знает о передаче невидимых, неосязаемых и неслышимых вибраций через электромагнитное поле, может отсюда заключить, что музыку можно полностью объяснить через составляющие радиоприемника, способ их расположения и энергию, от которой зависит их действие. Если же такой человек допускает возможность того, что нечто приходит в приемник извне, то он бы ее отверг, обнаружив, что приемник имеет одинаковый вес независимо от того, включен он или выключен. Таким образом, он должен был бы предположить, что ритмическая и гармоничная музыка возникает внутри радиоприемника вследствие невероятно сложных взаимодействий между его элементами. После тщательного изучения и анализа состава приемника он даже может быть способен создать его копию, производящую точно такие же звуки, как и оригинал, и, вероятно, может рассматривать такой результат как замечательное подтверждение своей теории. Но, несмотря на это свое достижение, он останется в полном неведении относительно того, что в действительности источником музыки была радиостанция на расстоянии в сотни миль. В рамках гипотезы формативной причинности «передача» приходит от предыдущих подобных систем, а ее «прием» зависит от деталей структуры и организации принимающей системы. Как и в радиоприемнике, существенное влияние здесь оказывают два вида изменений в организации «приемника». Во-первых, изменения в «настройке» системы могут приводить к приему совершенно различных «передач»: как радиоприемник можно настроить на разные радиостанции, так и развивающаяся система может быть «настроена» на разные морфогенетические поля. Во-вторых, как изменения в радиоприемнике, настроенном на определенную станцию, могут приводить к изменениям и искажениям музыки, исходящей из громкоговорителя, так же и изменения в системе, развивающейся под влиянием определенного морфогенетического поля, могут приводить к различным изменениям и искажениям конечной формы. Таким образом, в развивающихся организмах как окружающая среда, так и генетические факторы могут влиять на морфогенез двумя различными способами: либо через изменения «настройки» морфогенетических зародышей, либо изменяя обычные пути морфогенеза так, что образуются разные варианты нормальных конечных форм. 7.2. Измененные морфогенетические зародыши Морфогенетические зародыши для развития органов и тканей состоят из клеток или групп клеток с характерными структурой и организацией колебаний (разделы 4.5,6.1). Если в силу необычных условий окружающей среды или генетических изменений структура и организация колебаний зародыша достаточно изменились, то он перестает быть связанным с обычным для него морфогенетическим полем: тогда он либо может вообще стать неспособным действовать как зародыш, и в этом случае вся соответствующая структура не появится в организме; либо он может оказаться связанным с другим морфогенетическим полем, и тогда в этой части организма может развиться иная структура, а не та, которая обычно здесь образуется. Было описано множество примеров такой потери целостной структуры или замены одной структуры на другую. Иногда одинаковые изменения могут быть вызваны генетическими факторами и изменениями в окружении развивающегося организма; последние в генетической литературе именуются фенокопиями.[147] 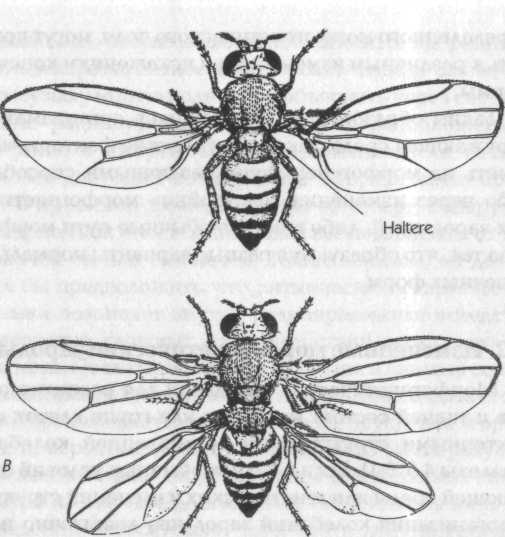 Рис. 17. Нормальная особь плодовой мушки Drosophila (А) и мутантная мушка (В), у которой третий грудной сегмент был трансформирован таким образом, что он стал похож на второй грудной сегмент. В результате мушка имеет две пары крыльев вместо одной Эффекты такого рода были детально изучены на плодовой мушке дрозофиле. Значительное число идентифицированных мутаций приводит к трансформациям целых участков тела мушки; например, антеннопедия заменяет усики на ножки, а мутации в грудном (бито-раксовом) генном комплексе приводит к тому, что третий (задний) грудной сегмент, который в норме содержит два зачатка (halteres), развивается так, как если бы он был средним сегментом (рис. 17). При этом взрослые организмы имеют две пары крыльев на соседних сегментах.[148] Сопоставимые явления были обнаружены у растений. Например, у гороха в норме листья включают собственно листочки у основания и усики на конце. В некоторых листьях усики образуются напротив листьев, указывая на то, что одинаковые примордии (зачаточные листы) могут давать структуры обоих типов (рис. 18); предположительно на клетки в этих примордиях влияют некие факторы в эмбриональном листе, заставляющие эти клетки принимать структуру и организацию колебаний, характерные для морфогенетического зародыша либо усика, либо листочка. Однако в мутанте одного типа способность образовывать усики подавлена, и все примордии дают листочки; в мутанте другого типа (вследствие гена в другой хромосоме) подавлено образование листочков, и все примордии дают только усики[149] (рис. 18). 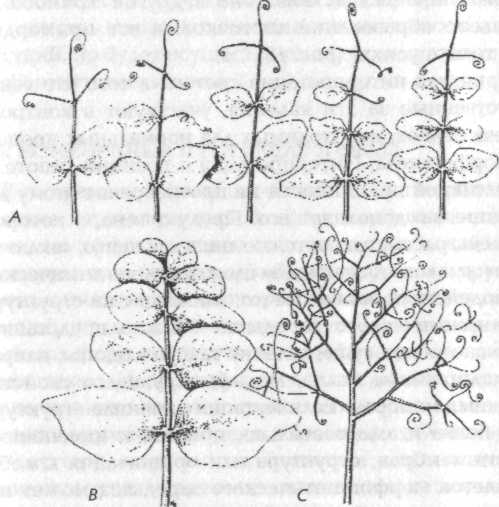 Рис. 18. А — нормальные листья гороха, имеющие как листочки, так и усики; В — лист мутанта, у которого образуются только листочки; С — лист мутанта, у которого образуются только усики Принятая интерпретация состоит в том, что гены, ответственные за эти явления, участвуют в контроле синтеза белков, необходимых для нормальных процессов морфогенеза. Интерпретация с позиции гипотезы формативной причинности не противоречит этому допущению, но дополняет его. Продукт гена, о котором идет речь, рассматривается не лишь как нечто, «включающее» или «выключающее» сложную цепь химических взаимодействий, но как нечто, влияющее на структуру морфогенетического зародыша. Можно представить много способов осуществления такого влияния, например кодирование белка, модифицирующего свойства клеточных мембран. Если мутация изменила структуру этого белка и, следовательно, привела к изменениям свойств мембран, структура или организация колебаний клеток морфогенетического зародыша может измениться таким образом, что они уже не могут быть связаны с обычным для них морфогенетическим полем. Следовательно, весь путь морфогенеза будет заблокирован. Поскольку клетки, участвующие в этом пути, более не претерпевают нормальное развитие и дифференциацию, они не будут синтезировать белки, характерные для этих процессов. А если морфогенетический зародыш изменился так, что он оказался связанным с другим морфогенетическим полем с помощью морфического резонанса, то развивающиеся клетки будут синтезировать белки, соответствующие другому морфогенетическому процессу. Таким образом, мутация, в результате которой один из путей морфогенеза оказался заблокированным или которая привела к другому пути, действительно изменяет продукт гена, косвенно контролирующий заданную схему синтеза белков, как и предполагает механистическая теория. Однако этот контроль не будет зависеть только от сложных химических взаимодействий, но будет осуществляться также посредством морфогенетических полей. 7.3. Измененные пути морфогенеза В то время как факторы, влияющие на морфогенетические зародыши, производят в морфогенезе качественные эффекты, такие как отсутствие какой-либо структуры или замена одной структуры на другую, многие генетические факторы или воздействия окружающей среды вызывают количественные изменения конечных форм посредством модификации процессов морфогенеза. Например, в растениях данной культурной разновидности, выросших в некотором диапазоне условий окружающей среды, строение стебля и корневой системы, морфология листьев и даже анатомия различных органов в деталях отличаются, но тем не менее характерная форма этой разновидности остается узнаваемой. Или в различных разновидностях данного вида, выросших в одинаковых условиях, растения отличаются друг от друга множеством деталей, хотя все они узнаваемы как разные варианты одной характерной формы: в конечном счете виды определяются прежде всего по их морфологии. Генетические факторы и окружающая среда влияют на развитие через различные количественные воздействия на компоненты структуры, ферментативную активность, гормоны и т. д. (раздел 7.1). Некоторые из этих воздействий сравнительно неспецифичны и изменяют несколько различных путей морфогенеза. Другие могут нарушать нормальный ход развития, но мало влияют на конечную форму благодаря регуляции. Хотя некоторые поразительные генетические эффекты и могут быть прослежены до конкретных генов, большая часть таких эффектов зависит от комбинированного воздействия множества генов, индивидуальные эффекты которых малы, их трудно выявить и подвергнуть анализу. Согласно гипотезе формативной причинности, организмы одного вида или сорта будут похожи друг на друга не только потому, что они генетически подобны и, следовательно, подвержены одинаковым генетическим влияниям в процессе морфогенеза, но также и потому, что характерные хреоды их вида усилены и стабилизированы посредством морфического резонанса от прошлых организмов того же вида. Морфогенетические поля видов не фиксированы, но изменяются с эволюцией вида. Наибольший статистический вклад в вероятностные структуры морфогенетических полей будет внесен наиболее общими морфологическими типами, которые одновременно будут теми, которые развивались при наиболее обычных условиях среды. В простейших случаях эффект автоматического усреднения при морфическом резонансе будет стабилизировать морфогенетические поля вокруг одиночных наиболее вероятных форм или «дикого типа». Но если вид обитает в двух географически или экологически различающихся местностях, в которых развивались характерные разновидности или сорта, морфогенетические поля видов будут содержать не единственную наиболее вероятную форму, а «мультимодальное» распределение форм, зависящее от числа морфологически различимых разновидностей или сортов и от относительной численности их прошлых популяций. 7.4. Доминирование На первый взгляд может показаться, что идея о стабилизации видовых форм путем морфического резонанса от прошлых организмов того же вида мало что добавляет к обычному объяснению в рамках одного лишь генетического подобия. Однако важность этой идеи становится очевидной при рассмотрении гибридных организмов, которые подвержены морфическому резонансу от двух различных родительских типов. 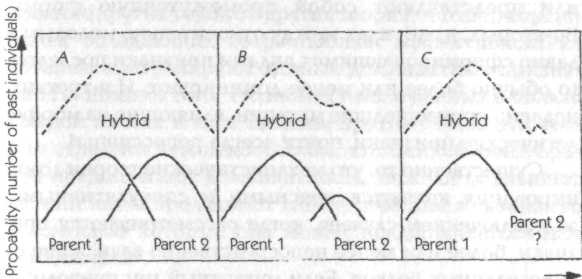 Рис. 19. Диаграмма вероятностных структур морфогенетических полей родителей и гибридов Вернемся к аналогии с радио: при нормальных обстоятельствах приемник настроен в любой момент времени только на одну станцию, так же как организм в норме «настроен» на прошлые подобные системы того же вида. Но если радио должно быть настроено на две станции одновременно, то звуки, которые оно издает, зависят от относительной силы сигналов этих станций: если один очень сильный, а второй очень слабый, то эффект последнего мало заметен; но если оба одинаково сильны, приемник издает смесь звуков из обоих источников. Подобно этому, в гибриде, образовавшемся в результате скрещивания двух видов, присутствие генов и характерных для каждого вида генных продуктов будет стремиться привести развивающийся организм в состояние морфического резонанса с прошлыми организмами обоих родительских типов. Тогда полный набор вероятностных структур в пределах морфогенетических полей гибрида будет зависеть от относительной силы морфического резонанса двух родительских типов. Если оба родителя происходят из видов, представленных сравнимым числом прошлых индивидуумов, то оба будут влиять на морфогенез в одинаковой степени, давая комбинацию или «результирующую» двух родительских форм (рис. 19 А). Но если один вид был представлен меньшим числом индивидов, чем другой, меньший вклад первого в общую вероятностную структуру будет означать, что станет преобладать форма второго из родительских видов (рис. 19 В). А если один из родителей происходит из недавно образовавшейся мутантной линии, морфический резонанс от малого числа прошлых индивидуумов этого типа внесет лишь незначительный вклад в вероятностную структуру гибридов (рис. 19 C). Эти ожидания согласуются с фактами. Во-первых, гибриды между двумя хорошо утвердившимися разновидностями или видами обычно сочетают черты обоих или представляют собой промежуточную форму. Во-вторых, в гибридах между относительно молодым и давно сформировавшимся видами признаки последнего обычно более или менее доминируют. И в-третьих, недавно произошедшие мутации, влияющие на морфологические признаки, почти всегда рецессивны.[150] Существенно то, что механистические теории доминирования являются неясными и спекулятивными, за исключением случаев, когда рассматриваются признаки, более или менее непосредственно зависящие от определенных белков. Если мутантный ген приводит к потере функции, например стимулируя синтез дефектного фермента, то он будет рецессивным, поскольку в гибридах присутствие нормального гена делает возможным производство нормального фермента и, следовательно, протекание нормальных биохимических реакций. Однако в некоторых случаях продукт дефектного гена может быть опасным, например нарушая проницаемость мембран, и тогда мутация будет одновременно доминирующей и летальной. Эти объяснения удовлетворительны в пределах своей применимости; но в отсутствие какого-либо механистического понимания морфогенеза попытка объяснить доминирование в наследовании формы путем экстраполяции с молекулярного уровня неизбежно приводит к необходимости считать этот спорный вопрос решенным и не требующим доказательств. Принятые генетические теории доминирования более сложные, нежели чисто биохимическая теория; они подчеркивают, что доминирование не фиксировано, но оно эволюционирует. Чтобы объяснить относительную однородность диких популяций, в которых большинство нелетальных мутаций рецессивны, они допускают, что доминированию «дикого типа» благоприятствует естественный отбор. Одна теория постулирует отбор генов, которые модифицируют доминирование других генов,[151] а другая теория — отбор версий генов, обладающих возрастающей эффективностью, которые контролируют данные доминантные признаки.[152] Но помимо того, что имеется мало данных в пользу каждой из них и есть таковые против обеих, эти теории страдают тем недостатком, что они, скорее, заранее предполагают доминантность, нежели объясняют ее: они лишь предлагают гипотетические механизмы, с помощью которых она может поддерживаться или усиливаться.[153] Согласно гипотезе формативной причинности, доминантность развивается по причине фундаментально иного характера. Типы, которым благоприятствует естественный отбор, будут представлены большим числом индивидуумов, чем типы с меньшей способностью к выживанию; с течением времени первые приобретают возрастающую доминантность в результате кумулятивного эффекта морфического резонанса. Эту гипотезу в принципе можно экспериментально отличить от всех механистических теорий доминантности. Согласно последним, при данных условиях окружающей среды доминантность зависит только от генетического строения гибрида, тогда как, согласно первой, она зависит как от генетического строения, так и от морфического резонанса родительских типов. Поэтому, если соотношение сил резонанса от родительских типов изменяется, то доминирование одного над другим также изменится, даже если генетическое строение гибрида останется тем же. Рассмотрим следующий эксперимент. Гибридные семена получены скрещиванием между растениями четко определенного сорта (Р1) и мутантной линии (Р2). Некоторые из этих гибридных семян помещены в холодный инкубатор, а другие выращиваются в контролируемых условиях. Характеристики гибридных растений тщательно регистрируются, и сами растения сохраняются. В этих растениях абсолютно доминирует морфология Р1 (рис. 19 С). Затем очень большие количества растений мутантного типа (Р2) выращиваются в поле. Некоторые из полученных затем гибридов снова выращиваются в тех же условиях, что и раньше, из той же партии семян. Поскольку теперь Р2 вносит больший вклад путем морфического резонанса, Р1 может доминировать лишь отчасти (рис. 19 В). После нового выращивания множества растений Р2 форма гибрида может стать промежуточной между двумя родительскими типами (рис. 19 А). Затем выращивается еще больше растений Р2; после этого гибриды снова выращиваются в тех же условиях, что и предыдущие из той же партии семян. Теперь тип Р2 будет вносить путем резонанса больший вклад, чем Р1 и морфология Р2 станет доминантной. Такой результат явится весомым свидетельством в пользу гипотезы о роли морфического резонанса в доминировании признаков и будет абсолютно непонятным с точки зрения ортодоксальной генетической теории. Единственная проблема в таком эксперименте состоит в том, что его может быть трудно осуществить на практике, поскольку если Р1 — это хорошо определенный тип, который уже существовал очень долгое время — в случае дикого типа, возможно, тысячи или даже миллионы лет, — то практически невозможно вырастить сравнимые количества типа Р2. Эксперимент может быть осуществим, только если P1 — это недавно появившийся сорт, у которого количество особей, выращенных в прошлом, было сравнительно невелико. 7.5. Семейные сходства В пределах данного вида организмы отличаются друг от друга всевозможными мелкими деталями. В скрещенной популяции каждая особь более или менее уникальна генетически и потому стремится следовать собственному пути развития под различными количественными влияниями своих генов. Более того, поскольку морфогенез зависит от влияния вероятностных структур на вероятностные события, процесс в целом становится до некоторой степени неопределенным. А еще и местные условия различаются. В результате действия всех этих факторов каждый индивидуум имеет характерную форму и вносит собственный уникальный вклад в возникающие затем морфогенетические поля. Вероятно, наиболее специфичным морфическим резонансом, действующим на определенный организм, является тот, который проистекает от близко связанных индивидуумов со сходным генетическим строением, что объясняет семейное сходство. Этот специфичный морфический резонанс накладывается на менее специфичный резонанс от многочисленных прошлых индивидуумов той же разновидности; а этот, в свою очередь, будет накладываться на общий фон морфического резонанса от всех прошлых представителей вида. В модели хреоды (рис. 5) наиболее специфичные морфические резонансы будут определять конкретные пути индивидуального морфогенеза, соответствующие руслу ручья, а менее специфичный морфический резонанс от предыдущих индивидуумов той же разновидности будет соответствовать дну небольшой долины. Хреоды различных разновидностей в пределах данного вида соответствуют незначительно отклоняющимся или параллельным долинам в пределах более широкой долины, представляющей хреоду вида в целом. 7.6. Эффекты окружения и морфический резонанс На формы организмов в различной степени влияют условия среды, в которой они развиваются. Согласно гипотезе формативной причинности, на них влияют также условия среды, в которых развивались предыдущие подобные организмы, поскольку формы этих организмов вносят вклад в морфогенетические поля путем морфического резонанса. Используя аналогию с радио, можно сказать, что музыка, выходящая из радиоприемника, испытывает на себе влияние изменений не только в приемнике, но и в передающей станции; если оркестр начинает играть другую музыку, радиоприемник также производит другие звуки, хотя его настройка и внутренняя структура не изменились. Рассмотрим, например, новую разновидность культурного вида. Если очень большие количества растений этого вида выращены в одной среде, а очень малые — в другой, то первые будут вносить существенно больший вклад в вероятностные структуры морфогенетических полей этой разновидности; их форма будет наиболее вероятной формой разновидности и потому будет влиять на морфогенез всех последующих растений той же разновидности, даже тогда когда они будут выращиваться в другой среде. Чтобы проверить это предсказание, вероятно, лучше всего было бы использовать новую разновидность самоопыленного зерна; такие растения очень близки друг другу генетически, и не будет опасности их скрещивания с другими разновидностями. Для начала несколько растений должны быть выращены в двух очень разных окружающих условиях, X и Y, и их морфологические признаки тщательно зарегистрированы. Некоторые из начальной партии семян должны быть заложены в холодное хранилище. Затем очень большие количества растений выращиваются в условиях Y (либо в один сезон, либо несколько поколений). Далее из нескольких семян, хранившихся на холоде, снова выращиваются несколько растений в условиях X. На их морфогенез теперь должен влиять морфический резонанс от больших количеств генетически подобных растений, выращенных в условиях Y. Следовательно, они должны иметь больше признаков морфологии Y-типа, чем имели первичные растения Х-типа. (Конечно, для корректного сравнения растений, выращенных в Х-условиях в различных случаях, необходимо было бы удостовериться, что условия были практически одинаковыми; это невозможно сделать в поле, но довольно легко обеспечить в искусственно контролируемой среде в фитотроне.) Если бы такой результат действительно был получен, он дал бы положительное свидетельство в пользу гипотезы формативной причинности и был бы необъясним с позиций механистической теории. Негативный результат здесь был бы неубедителен по двум причинам: во-первых, прямые воздействия условий X на морфогенетические процессы могут быть столь сильными, что они всегда будут канализировать морфогенез по хреодам Х-типа, несмотря на относительно слабый эффект морфического резонанса для этих путей. И во-вторых, растения других разновидностей того же вида будут влиять на развитие посредством морфического резонанса, хотя и менее специфично; тем не менее это влияние будет стремиться стабилизировать или хреоды обоих типов, X и Y, или каждую в отдельности, особенно если эти условия похожи на те, в которых обычно росли предыдущие разновидности того же вида. Этот эффект может быть минимизирован надлежащим выбором условий среды. 7.7. Наследование приобретенных признаков Влияние предшествующих организмов на последующие подобные организмы путем морфического резонанса может приводить к эффектам, которые невозможно было бы предположить в случае, если наследственность зависит только от передачи генов или других материальных структур от родителей к потомству. Такое обстоятельство позволяет увидеть в ином свете вопрос о наследовании приобретенных признаков. В упорном противостоянии друг другу в конце девятнадцатого — начале двадцатого столетия как ламаркисты, так и последователи Вейсмана и Менделя полагали, что наследственность зависит только от зародышевой плазмы вообще или от генов в частности. Поэтому, если признаки, приобретенные организмом в ответ на воздействие среды, должны были наследоваться, то или зародышевая плазма, или гены должны были претерпевать специфические изменения. Противники ламаркизма подчеркивали, что такие изменения представляются чрезвычайно маловероятными, если не невозможными. Даже сами ламаркисты были неспособны предложить какие-либо правдоподобные механизмы, которые могли бы привести к таким изменениям. С другой стороны, теория Ламарка, казалось, давала правдоподобные объяснения наследственным адаптациям у животных и растений. Например, у верблюдов на коленях есть мозоли. Легко понять, что они приобретаются в результаты стирания кожи, когда верблюды становятся на колени. Но верблюжата рождаются уже с мозолями. Факты такого рода могут стать вполне понятными, если приобретенные признаки каким-то образом наследуются. Однако менделисты такую возможность отрицают и предлагают альтернативную интерпретацию, используя идею о случайных мутациях: если организмы с данными приобретенными признаками являются более приспособленными с точки зрения естественного отбора, то случайные мутации, которые приводят к появлению таких же признаков без необходимости их наследования, также будут усиливаться путем естественного отбора, и, таким образом, эти признаки станут наследуемыми. Такое гипотетическое представление наследования приобретенных признаков иногда называют эффектом Болдуина — по имени теоретика-эволюциониста, который впервые его предложил.[154] В начале двадцатого столетия многие ученые утверждали, что им удалось продемонстрировать наследование приобретенных признаков у различных видов животных и растений.[155] Антиламаркисты ответили контрпримерами, снова и снова цитируя хорошо известный эксперимент Вейсмана, в котором он отрубал хвосты у мышей в двадцати двух последовательных поколениях и обнаружил, что их потомство все равно рождается с хвостами. Другой аргумент привлекал внимание к тому факту, что после обрезания у многих поколений евреев они все еще рождаются с крайней плотью. После самоубийства одного из ведущих ламаркистов, П. Каммерера, в 1926 году менделизм утвердился на Западе как почти неоспоримая общепринятая теория.[156] Между тем в Советском Союзе последователи идеи о наследовании приобретенных признаков во главе с Т. Д. Лысенко заняли ведущие позиции в руководстве биологической наукой в 1930-х гг. и сохраняли их до 1964 г. В этот период многие из их оппонентов — менделистов — подверглись жестоким гонениям.[157] Такая поляризация породила горечь и догматизм в обоих направлениях. Однако сейчас имеется достаточно много свидетельств того, что приобретенные признаки действительно могут наследоваться; проблема перешла в область интерпретации. В своих выдающихся сериях экспериментов К. X. Уоддингтон подвергал яйца или куколки дикой популяции плодовой мушки воздействию либо паров эфира, либо высоких температур, вызывая тем самым ненормальное развитие некоторых мушек.[158] Следующее поколение выводилось от этих ненормальных мушек, яйца или куколки снова подвергались стрессовым воздействиям среды; и снова отбирались ненормальные мушки, от них получали потомство и так далее. В последовательных поколениях процент ненормальных мушек возрастал. Через много поколений (в некоторых случаях 14, в других — 20 или более), когда потомство ненормальных мушек выращивалось без стрессовых воздействий, в нормальных условиях среды некоторые из них все еще развивались с характерными отклонениями. Более того, эти отклонения продолжали появляться в их потомстве, выращенном в нормальных условиях. По словам Уоддингтона: «Все эти эксперименты показывают, что если отбор происходит по наличию признака, приобретенного в данных ненормальных условиях окружающей среды, полученные избранные линии могут проявлять этот признак даже тогда, когда они возвращаются к нормальному окружению».[159] Уоддингтон рассматривал возможность того, что некоторые физические или химические воздействия от измененных структур в ненормальных мушках могли вызывать наследуемые модификации в их генах,[160] но отверг ее, поскольку открытия молекулярной биологии показали, что любой из таких механизмов крайне маловероятен.[161] Его заключительная интерпретация подчеркивала как роль отбора для отклика генетического потенциала на стрессовые воздействия путем ненормального развития, так и «канализацию развития», происходящую при модифицированном морфогенезе. «Используя несколько образный язык, можно сказать, что отбор не просто понизил порог, но и определил, в каком направлении будет двигаться развивающаяся система, когда она этот порог перейдет».[162] Сам Уоддингтон создал слово «хреода» для выражения идеи направленного, канализированного развития. Он думал об определении направления, принимаемого хреодой, в терминах ее «настройки». Но он не объяснил, как возникают эта канализация и «настройка», сделав лишь неопределенное предположение, что они каким-то образом зависят от отбора генов.[163] Гипотеза формативной причинности дополняет интерпретацию Уоддингтона: хреоды и конечные формы, к которым они направлены, зависят от морфического резонанса от предыдущих подобных организмов; наследование приобретенных признаков типа, исследовавшегося Уоддингтоном, зависит как от генетического отбора, так и от прямого влияния морфического резонанса от организмов, развитие которых было модифицировано в ответ на ненормальное окружение. Примечания:1 Особенно четкое представление дано у Monod (1972). 14 От оцифровщика. Отличия от оригинала: 1) Концевые сноски (примечания) преобразованы в обычные (перемещены на текущую страницу); 2) Перемещено с конца книги в начало «Содержание» 15 Huxley (1867), р. 74. 16 Витализм (от лат. Vita — жизнь) — учение о качественном отличии живой природы от неживой, о принципиальной несводимости жизненных процессов к законам и силам неорганического мира, о наличии в живых телах особых факторов, отсутствующих в неживых. — Прим. пер. 147 Или организмами (индивидуумами), которые при непосредственном наблюдении демонстрируют одинаковый набор внешних признаков, то есть относятся к одному фенотипу. Фенотип (от греч. jainw — показываю, обнаруживаю) — совокупность свойств и признаков организма, сложившихся в процессе его индивидуального развития (онтогенеза). Фенотип определяется наследственной основой (генной конституцией, или генотипом) и условиями среды, в которой протекает онтогенез. — Прим. пер. 148 Morata and Lawrence (1972). 149 Snoad (1974). 150 То есть у потомства выражены слабо или отсутствуют, в противоположность доминантным признакам, которые преобладают или полностью вытесняют аналогичный признак другого родителя. Соответственно, ген, ответственный за существование данного признака у одного из родителей, является доминантным, а у другого — рецессивным. Явление доминирования и закон расщепления признаков, хромосомная и генетическая теории наследственности, роль мутаций освещаются в специальной литературе (например, Дж. Уотсон. Молекулярная биология гена. М.: Мир, 1967; Н. Грин, У. Стаут, Д. Тейлор. Биология. В 3 томах. М.: Мир, 1993. Т. 3, гл. 22, 23). — Прим. пер. 151 Fisher (1930). 152 Haldane (1939). 153 Serra(1966). 154 Boldwin (1902). 155 Многие из этих фактов собраны в Semon (1912) и Kamerer (1924). 156 Koestler(1971). 157 Medvedev(1969). 158 Описания этих экспериментов удобно собраны в Waddington(1975). 159 Там же, с. 65. 160 Waddngton(1957). 161 См. дискуссию между Уоддингтоном и Кестлером в Koestler and Smythies (eds) (1969), с. 382–391. 162 Waddingon(1975),c. 87. 163 Там же, с. 87–88. |
|
||
|
Главная | В избранное | Наш E-MAIL | Прислать материал | Нашёл ошибку | Верх |
||||
|
|
||||